Jedinečnost intracelulární struktury červených řas spočívá jak ve vlastnostech běžných buněčných složek, tak v přítomnosti specifických intracelulárních inkluzí.
Buněčné membrány. V buněčných stěnách červených řas jsou jasně rozlišeny dvě vrstvy: vnitřní, sestávající z celulózy, a vnější, tvořená pektinovými sloučeninami. U forem s hrubým, tvrdým stélkem se mimo buněčné stěny vyvíjí tenká vrstva kutikuly. Složením a strukturou se kutikula šarlatových rostlin liší od kutikuly vyšších rostlin. Například v porfyru vzniká jako výsledek kondenzace molekul monosacharidu manózy. U měkkých slizničních forem kutikula chybí.
Pektinové látky v červených řasách jsou vápenaté a hořečnaté soli speciálních pektinových kyselin. Mají schopnost rozpouštět se ve vroucí vodě za vzniku slizových roztoků. Do skupiny pektinových látek patří i speciální koloidní látky, které jsou obsaženy v buněčných membránách a mezibuněčných prostorech mnoha šarlatových hub. Jsou komplexní směsí polysacharidů obsahujících síru a souhrnně se nazývají fykokoloidy. Fykokoloidy se nerozpouštějí ve studené vodě, ale dobře se rozpouštějí ve vroucí vodě za vzniku koloidních systémů. V současnosti jsou nejdůležitějšími produkty získávanými z mořských řas fykokoloidy červených řas a také hnědé řasy. Vzhledem k tomu, že tyto látky jsou široce používány v lidském ekonomickém životě, jejich chemické složení bylo poměrně dobře prozkoumáno. Fykokoloidy se získávají z mnoha druhů a v důsledku toho bylo založeno několik odrůd. Nejznámější jsou agar, karagenan, nori a agaroidy. Tyto látky se od sebe liší složením a vlastnostmi, ale mají společnou gelující schopnost.
Přítomnost koloidních látek určuje schopnost buněčných membrán po smrti silně bobtnat, což ztěžuje jejich obarvení pro zkoumání.
Mezi červenými řasami existují formy, jejichž schránky jsou zvápenatělé. Nejprve se ve střední desce, mezi vnější a vnitřní vrstvu pláště, uloží vápno, které postupně proniká celulózovou vrstvou a více či méně silně ji impregnuje. Ale i při silném zavápnění zůstává uvnitř vždy tenká membrána bez vápna, která odděluje plazmu od vápenaté vrstvy.
Složení vápenných sloučenin není stejné. Kalcit se ukládá u korálovitých (corallineaceae) a aragonit u některých jiných než malinovitých. Kromě toho se nacházejí uhličitany vápenaté a hořečnaté a také železo.
Růst skořápek probíhá následovně. Jeho nové vrstvy jsou položeny na vrcholu protoplastu apikální buňky, zatímco vnější vrstvy se postupně oddělují, jak buňka roste. Výsledkem je, že celá skořápka má vrstvenou strukturu, a protože nové vrstvy jsou uloženy pod určitým úhlem, vypadá jako trychtýř.
Cytoplazma červených řas se vyznačuje vysokou viskozitou a pevně přilne k buněčným stěnám. Velmi snadno podléhá plazmolýze, a proto jsou červené řasy velmi citlivé na změny podmínek prostředí, zejména na odsolování.
Jádro. Mezi červenými řasami existuje velké množství forem s mononukleárními buňkami. Zpravidla se jedná o nejjednodušší organizované formy. U vysoce organizovaných šarlatových buněk jsou buňky většinou vícejaderné, výjimkou jsou mladé buňky stélku – apikální, jádrové atd. Mononukleární buňky větví omezeného růstu za určitých podmínek se stávají vícejadernými. Lze však najít i řasy, u kterých jsou staré buňky centrálních filament jednojaderné a mladší buňky je obklopující naopak obsahují několik jader. Reprodukční buňky – spermie, karpospory, tetraspory – mají vždy jedno jádro, ale vajíčka jsou často obklopena vícejadernými buňkami, a to i u forem, jejichž celý stélek se skládá z mononukleárních buněk. Jádro červené řasy je malé, má čirou jadernou membránu a jadérko.
Chloroplasty. Buňky červených řas obsahují jeden nebo více chloroplastů. Rostliny třídy Bangiaceae a primitivní zástupci nejnižšího organizovaného řádu Nemaliaceae mají jeden hvězdicovitý chloroplast s jedním pyrenoidem. Obvykle zaujímá v buňce axiální polohu a skládá se pak z centrálního tělesa a výběžků z něj vybíhajících do všech směrů (obr. 149, 2). V axiálním chloroplastu se pyrenoid nachází v jeho středu. Procesy vycházející z centrálního chloroplastu se mohou rozšířit na periferii a blízko u sebe, čímž se vytvoří nepravidelný nebo páskovitý stěnový chloroplast. Pravděpodobně je stěnový chloroplast většiny fialových chloroplastů odvozen od axiálního chloroplastu v důsledku ztráty centrální části.

Role pyrenoidu v červených řasách není příliš jasná. V některých případech je jeho přítomnost spojena s ukládáním škrobových zrn; v jiných se pyrenoid nachází v buňkách, které se neúčastní asimilačních procesů. Ve více organizovaných formách pyrenoid mizí; tento proces lze vysledovat již u řádu nemaliaceae.
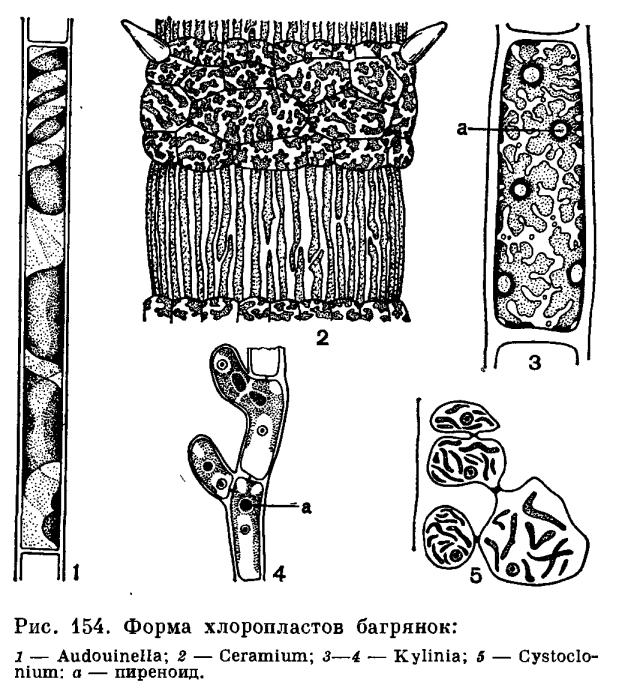
U červených řas postrádajících pyrenoidy jsou chloroplasty dvou hlavních typů – páskovité a čočkovité (nebo diskovité) s četnými přechody mezi nimi (obr. 154). Druhy na nižším stupni evoluce mají obvykle stuhovitý chloroplast; pro vysoce organizované formy jsou naopak charakteristické chloroplasty čočkovitého tvaru. Totéž lze říci o počtu chloroplastů – jejich počet se zvyšuje se složitostí organizace. Tvar chloroplastu není něco stálého, může se měnit s věkem, se světelnými podmínkami, se změnami velikosti buněk, i když velké skupiny šarlatových rostlin se často vyznačují chloroplastem určitého tvaru. U Ceramia jsou chloroplasty ve velkých buňkách internodií protáhlé, stuhovité a v krátkých buňkách jádra v uzlinách jsou to krátké, nepravidelně laločnaté destičky. Reprodukce chloroplastů probíhá jednoduchým propletením, jako u vyšších rostlin.
Ve své jemné struktuře viditelné pouze pod elektronovým mikroskopem se chloroplasty červených řas téměř neliší od chloroplastů jiných řas.
Pigmenty. Šarlatové květy se vyznačují komplexní sadou pigmentů. Kromě v alkoholu rozpustného chlorofylu, karotenu a xantofylu běžných pro zelené rostliny obsahují chloroplasty červených řas další ve vodě rozpustné pigmenty, biliproteiny. Jedná se o fykoerythrin a fykocyanin. Zelené rostliny, jak řasy, tak suchozemské, obsahují dvě modifikace chlorofylu – modrozelený chlorofyl a a žlutozelený chlorofyl b. V červených řasách se nachází pouze chlorofyl a – univerzální pigment charakteristický pro všechny rostliny. V některých šarlatových rostlinách byl navíc nalezen chlorofyl d, jehož povaha však zůstává nejasná. Fialové rostliny obsahují ve srovnání s vyššími rostlinami málo zelených pigmentů a jsou obvykle maskovány dalšími biliproteiny. Určitý vzorec byl zaznamenán ve změně množství chlorofylu v závislosti na množství světla. Řasy přizpůsobené životu v podmínkách slabého osvětlení polárních moří jsou obvykle bohatší na chlorofyl než řasy z jižních moří. Stejně tak jsou hlubinné řasy bohatší na chlorofyl než ty, které rostou blízko hladiny vody.
Karotenoidy červených řas jsou zastoupeny α- a β-karotenem a xantofyly lutein, zeaxanthin a pravděpodobně i taraxanthin. Biliproteiny šarlatových hub jsou červený fykoerythrin a modrý fykocyanin. Jsou blízké pigmentům modrozelených řas, ale nejsou s nimi totožné, protože se liší chemickým složením. Jak bylo prokázáno v četných experimentech, počet pigmentů v šarlatových houbách roste s hloubkou; v tomto případě se množství fykoerythrinu zvyšuje ve větší míře než množství chlorofylu. Kdo tyto řasy sbíral v přírodě, ví, že červeně zbarvené řasy rostou v hloubce a že v mělké vodě mění barvu. S přibývajícím množstvím světla se stávají bledě červenými, poté žlutozelenými, slámově zbarvenými a nakonec zcela vybělené.
Existuje teorie tzv. chromatické adaptace, podle které je pronikání řas do určitých hloubek spojeno s kvalitou světla procházejícího vodním sloupcem. Jak víte, paprsky ze zelené a modré části spektra pronikají nejhlouběji. Červené pigmenty šarlatových řas jim umožňují fotosyntézu v modrém světle, a proto podle této teorie pronikají do hloubek nepřístupných jiným řasám. V praxi však tento vzorec není vždy dodržován. Jaká je role biliproteinů ve fotosyntéze červených řas? V experimentech bylo zjištěno, že při slabém osvětlení se podílejí na zvýšené absorpci světla. Proto je lze považovat za optické senzibilizátory. Pronikání červených řas do významných hloubek je tedy správněji vysvětleno jejich schopností absorbovat malé množství světla. Obecně platí, že fialové řasy jsou organismy odolné vůči stínu: jsou schopny lépe využívat slabé světlo než jiné řasy. Jestliže červená barva řas ve slabém světle prospívá, pak v silnějším světle je naopak intenzita fotosyntézy šarlatových řas nižší než u jiných řas, právě kvůli přítomnosti červených pigmentů. K ochraně před silným světlem mají šarlatové ryby žijící v mělkých hloubkách, zejména v tropických a subtropických mořích, zvláštní duhová těla. Tato matně žlutá tělíska nepravidelného tvaru se tvoří ve vakuolách povrchových buněk talu a sestávají z malých zrn bílkovinné povahy. Mají schopnost rozptylovat a odrážet sluneční paprsky dopadající na ně. Při velmi silném osvětlení jsou iridující tělíska umístěna pod vnější stěnou buňky, zatímco chloroplast je umístěn na vnitřní nebo boční stěně a slouží jako jakýsi závěs pro chloroplast. Když rostlina vstoupí do difuzních světelných podmínek, dojde k vzájemnému pohybu a chloroplast skončí u vnější stěny.
Řasy s duhovými těly mají v dopadajícím světle obvykle namodralý ocelový lesk. U některých druhů se v buňkách objevují velká čočkovitá tělesa, která s ubývajícím osvětlením mizí.
Náhradní látky. Jako produkt asimilace ukládají červené řasy polysacharid zvaný šarlatový škrob. Chemickou povahou je nejblíže amylopektinu a glykogenu a zjevně zaujímá mezipolohu mezi běžným škrobem a glykogenem. Fialový škrob se ukládá ve formě malých polotuhých tělísek různých tvarů a barev. Tato tělesa mohou mít tvar kuželů nebo plochých oválných desek s prohlubní na široké ploše. Často na nich můžete vidět soustředné zóny. Fialová škrobová zrna se tvoří částečně v cytoplazmě, částečně na povrchu chloroplastů, ale nikdy nevznikají uvnitř plastidů, na rozdíl od běžného škrobu zelených rostlin. Ve formách, které mají pyrenoid, se tento do určité míry podílí na syntéze škrobu.
Kromě šarlatového škrobu se jako rezervní látky v červených řasách ukládají cukry trehalóza, floridosid, sacharóza aj. V některých formách se hojně vyskytují vícemocné alkoholy. Z tuků jsou známy cholesterol, silosterol a fukosterol. Obsah tuku se liší v závislosti na podmínkách prostředí.
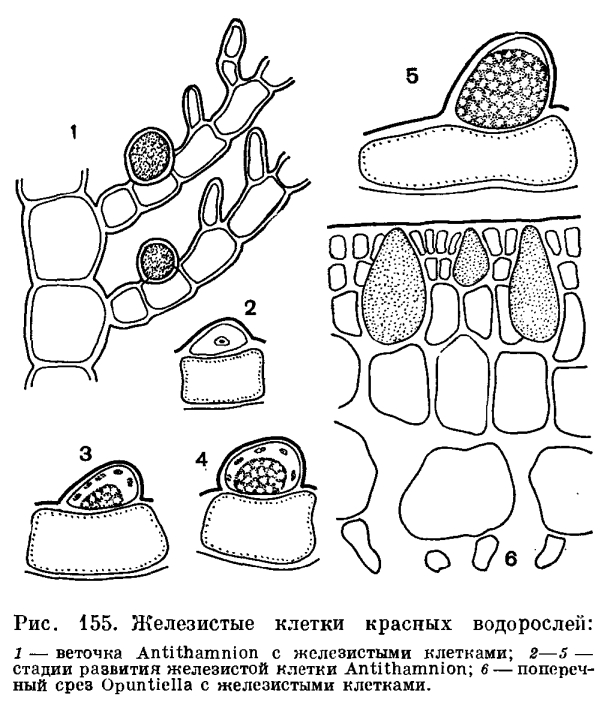
Žlázové buňky. Zvláštností červených řas je přítomnost speciálních buněk u některých zástupců floridské třídy s bezbarvými obsahy, které silně lámou světlo (obr. 155). V literatuře jsou známy jako vezikulární nebo žlázové buňky. Obsah těchto buněk je u různých řas různé povahy; jsou naplněny jodidovými, méně často bromidovými sloučeninami. Nejčastěji se žlázové buňky nacházejí v řádu ceramidů. Ve filamentózních rozvětvených řasách antitamnion sedí na horní straně postranních větví (obr. 155, 2). Při jejich vývoji se nejprve oddělí malá buňka čočkovitého tvaru obsahující malé množství plazmy a malé červené chloroplasty. Jádro lze vysledovat pouze v nejranější fázi vývoje. Brzy se na dně této buňky vytvoří bezbarvá bublina lámající světlo. Roste a spolu s ním se zvětšuje i celá buňka. Ve vzniklé buňce je většinu obsazena bublinou a pouze v horní části zůstává úzká vrstva plazmy s drobnými chloroplasty (obr. 155, 2-5). Role vezikulárních buněk nebyla objasněna, ačkoli v této záležitosti existuje mnoho různých předpokladů. Jsou považovány za nedostatečně vyvinuté sporangie, zásoby rezervních látek, „plavecké měchýře“ atd.
Žlázové buňky jsou charakteristické pro určité skupiny řas, a proto slouží jako důležitý taxonomický znak.
Vlasy. Tvorba chlupů je rozšířeným jevem ve třídě Florididae. Pravé šarlatové chlupy by měly být odlišeny od vlasových větví nebo falešných chlupů. U druhů s jednořadým vláknitým stélkem je vidět, jak se koncové buňky postranních větví prodlužují a odbarvují a získávají vlasovou strukturu (obr. 151, 7). To jsou falešné vlasy. Pravé chloupky červených řas se dělí na dva typy: jednobuněčné a mnohobuněčné. Jednobuněčné chlupy se nikdy nevětví. V jednořadých vláknitých formách se tvoří z apikálních buněk větví, u víceřadých – z povrchových buněk kůry. Buňka budoucího vlasu je od mateřské buňky oddělena příčnou přepážkou a je značně protáhlá, často dosahuje délky milimetru i více (obr. 156). Obsahuje jádro a malé množství cytoplazmy. Chloroplast mizí, jak vlasy rostou, a vlasy se stávají bezbarvými. Obvykle se mateřská buňka vlasu neliší od sousedních vegetativních buněk, ale někdy je mnohem větší a zůstává jasně viditelná i po opadnutí vlasu. V korálových vlasech nejsou chloupky samostatnými buňkami, ale jsou to pouze výrůstky speciálních buněk, od kterých nejsou odděleny přepážkou. Tyto buňky jsou mnohem větší než ostatní a jsou známé jako trichocyty nebo heterocysty (obr. 172, 2, 3). Pokud se ve třídě Floridaeidae vyskytují jednobuněčné chlupy poměrně často, pak u Bangiaceae zcela chybí.
Mnohobuněčné chlupy jsou obvykle více či méně silně rozvětvené. Nacházejí se pouze v některých řasách z řádu Ceramiaceae. Jak se ukázalo v experimentech, hlavní úlohou chloupků je to, že usnadňují vstřebávání živin z okolního prostředí.
Póry. Jednou z nejzajímavějších vlastností červených řas je to, že buňky, které tvoří stélku, jsou navzájem spojeny pomocí speciálních útvarů zvaných nory. Mezi dceřinými buňkami, tedy buňkami pocházejícími z jedné matky, se spojení provádí primárními póry (obr. 157, 2-3). Vznikají jako výsledek neúplné přepážky mezi dvěma nově vzniklými buňkami. Primární póry se nacházejí uprostřed septa, v místě, kterým lze nakreslit čáru spojující jádra dceřiných buněk, a představují tenkou desku. Řetězce procházejí touto destičkou a spojují cytoplazmu sousedních buněk. Až donedávna se věřilo, že primární póry jsou charakteristické pouze pro Floridaeidae a ve třídě Bangiaceae chybí – to byla jedna ze základních charakteristik, na základě kterých byly obě tyto třídy rozlišovány. V poslední době se však primární póry otevřely také u zástupců třídy Bangiaceae.

Kromě spojení dceřiných buněk dochází u většiny Floridaidae k sekundárnímu spojení sousedních buněk. Mohou to být buňky stejného vlákna, sousední buňky sousedních vláken nebo, u parazitických červů šarlatových, buňky parazita a hostitele. Více organizovaní zástupci třídy mají schopnost sekundárního spojení. Chybí v řádu Nemaliaceae a v primitivních řasách jiných řádů. Spojení sousedních buněk, mezi nimiž zpočátku nebylo žádné spojení, je provedeno sekundárními póry (obr. 157, 4-8). Tvorba sekundárních pórů je poměrně složitý proces. Začíná to tím, že se jádro v buňce rozdělí a jedno z dceřiných jader se přesune na periferii buňky. V místě vstupu jádra se vytvoří malý výrůstek, který se následně oddělí, jako malá buňka spojená s první přes primární pór. Roztažením se tato další buňka dostane k sousední a splyne s ní. Nyní se primární pór spojující další článek s prvním stane sekundárním. Někdy jsou dvě sousední buňky spojeny několika sekundárními póry. Role těchto sloučenin není plně pochopena; pravděpodobně usnadňují přenos živin z jedné buňky do druhé. I když je způsob tvorby primárních a sekundárních pórů odlišný, jejich struktura je podobná. A přesto ne všechny fialové ženy mají stejné póry. V některých případech existuje přímé cytoplazmatické spojení mezi buňkami, v jiných je mnohem méně zřejmé.
Životnost rostlin: v 6 svazcích. — M.: Osvěta. Za redakce A. L. Takhtadzhyana, šéfredaktora kor. Akademie věd SSSR, prof. A.A. Fedorov. 1974
Červené řasy neboli Rhabophyta (fialové řasy) jsou jednou z nejstarších skupin eukaryotických řas. Většině druhů dominují jednobuněčné řasy, mnoho známých druhů je mnohobuněčných.
Kromě dvou druhů nepohlavních tříd Cyanidiopyceae žijících v pobřežních jeskyních se již nevyskytují suchozemští zástupci červených řas. To může být způsobeno evolučním úzkým hrdlem, což znamená, že poslední společný předek ztratil téměř čtvrtinu svých esenciálních genů a většinu své evoluční flexibility (schopnosti se měnit). Červené řasy jsou v mořských biotopech rozmanité. Lze je nalézt i ve sladké vodě, ale ne tak často. Pouze asi 5 % červených řas se nachází ve sladkovodním prostředí. Vyšší koncentrace těchto řas se obvykle vyskytují v teplejších oblastech. Červené mořské řasy se nacházejí ve všech pobřežních mořských oblastech naší planety, ale jejich rozmanitost v chladných oblastech je mnohem menší než v teplých vodách. Červené řasy vedou většinou připoutaný životní styl – přichytávají se na kameny a mořské rostliny, mušle a korály. Speciální buňky červených řas pronikají do těla, na kterém se uchytí. Řasy jsou schopny vytvářet sekundární vazby s hostitelskými buňkami. Fialové řasy parazitují mimo jiné na dalších červených řasách.
Popis, charakteristické rysy a vlastnosti
- přítomnost eukaryotických buněk bez bičíků a centriol;
- přítomnost chloroplastů, ve kterých není vnější endoplazmatické retikulum.
Charakteristickým rysem červených řas je také použití fykobiliproteinů jako pomocných pigmentů, které jim dodávají hnědou nebo karmínovou barvu. Ačkoli červená barva šarlat je způsobena přítomností speciální látky v nich – fykoerythrinu, často mají jinou barvu: fialovou, nahnědlou kvůli dodatečné přítomnosti pigmentu fykocyaninu.
Tento typ řas má také další karotenoidní pigmenty, jejichž celkové množství závisí na hloubce přirozeného prostředí řas. Ve fialové skupině jsou také zelené řasy.
Řasy nacházející se výše ve vodním sloupci mají zpravidla širokou škálu pigmentů vhodných pro život v poměrně jasných podmínkách povrchových vod.
Řasy žijící v hloubce mají sadu dalších pigmentů. Tato sada obsahuje větší množství fykobilinů, takže řasy v hloubce jsou lépe přizpůsobeny fotosyntéze v podmínkách charakterizovaných nedostatkem světla.
Rezervní materiály červených řas jsou: polysacharidy červených řas, floridosidy a lipidové látky. Produkty fotosyntézy červených řas se hromadí ve formě fialových granulí škrobu. Červené řasy jsou schopny ukládat cukry ve formě škrobu jako zásoby potravy mimo své plastidy.
Fialový škrob má hnědou barvu díky obsahu jódu. Červené řasy obsahují látky obsahující cukry zvané floridosid a isofloridosid. Metabolismus sacharidů červených řas je velmi odlišný od metabolismu sacharidů skutečných rostlin.
Buněčná stěna se skládá ze dvou vrstev, vnitřní celulózy a vnějšího pektinu, který je nasycený látkami zabraňujícími vysychání, jako jsou: kyselina alginová, galaktóza: karagenan, xylan, mannan, agar, porfyr a furzel.
Plastidy červených řas mají různé tvary. Nejčastěji jsou plastidy obklopeny obalem sestávajícím ze dvou membrán a obsahují jednotlivé tylakoidy.
Plastidy červených řas obsahující jednotlivé tylakoidy jsou podobné jako u sinic. Liší se však tím, že jsou od sebe oddělené.
Thylakoidy jsou typem váčku uvnitř chloroplastů a sinic. Chloroplastové tylakoidy mají často strukturu podobnou hromadě disků.
Nejstarší známá červená řasa (stejně jako nejstarší známá eukaryota a pohlavně se rozmnožující organismus) je proterozoická Bangiomorpha pubescens, asi před 1,2 miliardami let.
Zástupci červených řas:
- Chondrus crispus neboli irský mech;
- polypsifonie;
- vidlička (Furcellaria);
- druh používaný v čínské a japonské kuchyni: jemný šeřík (Porphyra tenera).
Červené řasy, jako je palmaria a nori, jsou přísadami do různých asijských jídel a používají se v potravinářských výrobcích a doplňcích stravy (jako je agar, karagenan).
Druhy a klasifikace červených řas
V současné době je popsáno více než sedm tisíc druhů červených řas. K popisu nových druhů dochází neustále, každý rok přibývají nové druhy červených řas.
Většina červených řas žije v mořích, mnohem méně žije ve sladké vodě.
Klasifikace červených řas
Oddělení červených řas
Ve staré klasifikaci existují dvě třídy:
- bangi (Bangiophyceae);
- červené řasy (Florideophyceae).
Nová klasifikace zahrnuje rozdělení skupiny červených řas do tří tříd: první dvě podle staré klasifikace a třetí třída – cyanidium (Cyanidiophyceae).
Zvažte třídu červených řas Rhodophyceae, která má dvě podtřídy:
- Bangiophycidae;
- Florideophycidae.
Sbírka blízce příbuzných čeledí řas se v botanice nazývá řád. V zoologické klasifikaci pořadí odpovídá pořadí. Řády se někdy dělí na menší celky – podřády.
Třída Bangui (Bangiophyceae) se dělí do řádů:
- Bangiales;
- Cyanidiales;
- Erythropeltidales;
- Porphyridiales;
- Rhodochaetales;
- Stylonematales.
Podle novějších systémů založených na molekulárních fylogenetických výsledcích by měly být vyloučeny všechny řády kromě Bangiales. V tomto novém přístupu jsou Bangiophyceae monotypickou třídou. V některých systémech jsou červené řasy považovány za monotypickou skupinu s jedinou třídou Rhodophyceae.
Třída červených řas Florideophyceae se dělí na:
- Hildenbrandiophycidae;
- Nemaliophycidae;
- Corallinophycidae;
- Ahnfeltiophycidae;
- Rhodymeniophycidae.
Životní cyklus a reprodukce
Životní cykly červených řas jsou složité a ve většině případů se výrazně liší od ostatních skupin řas.
U rostlin probíhá obměna generací obvykle podle následujícího cyklu: g – c – g – c – g – c – g – c atd., kde g je gametofyt a c je sporofyt. Jedinou výjimkou jsou červené řasy, protože jejich transformace probíhá podle následujícího vzorce: g – c – c – g – c – c – g.
Jak se rozmnožují červené řasy
Většina červených řas je mnohobuněčných, makroskopických, mořských a rozmnožují se pohlavně. Jedná se o červené řasy, které vylučují uhličitan vápenatý. Podílejí se na výstavbě korálových útesů.
Během sexuální reprodukce produkuje haploidní gametofyt, nezávislá haploidní rostlina s karpogonálními buňkami vejčitého tvaru s karposporami a spermatangiemi, spermie.
Asexuální metoda reprodukce zahrnuje nepohyblivé spory, které postrádají bičíky. Existují třídy červených řas, které se rozmnožují vegetativně.
Celý životní cyklus zahrnuje tři generace řas:
- haploidní gametofyt, na kterém se gametangia nacházejí: samčí – spermatangia; samice – carpogonia;
- diploidní karposporofyt, vycházející ze zygoty a vyvíjející se na gametofytu a tvořící spory diploidního karpofytu;
- diploidní tetrasporofyt vznikající z klíčících karpospor, který v důsledku meiózy vytváří haploidní spory tetraspor, z nichž se tvoří haploidní gametofyty.
Nejčastěji postupuje gametofyt a tetrasporofyt stejně, ale jejich vzhled může být odlišný. Kvůli tomu byli rozděleni do různých rodů a dokonce do řádů vzdálených od sebe.
Struktura červených řas
Morfologie červených řas
Existují jednoduché jednobuněčné formy červených řas, stejně jako složité parenchymatické a neparenchymatické formy.
Červené řasy mají dvojité buněčné stěny. Vnější stěny obsahují polysacharidy agarózu a agaropektin, které lze z buněčných stěn extrahovat zahřátím. Vnitřní vrstvy obsahují převážně celulózu.
Po celý život nemají červené řasy bičíky a centrioly. Charakteristické rysy buněčných struktur červených řas jsou:
- normální vřetenová vlákna;
- mikrotubuly;
- fotosyntetické membrány;
- granule fykobilinového pigmentu;
- důlkové spojení.
Buňky červených řas nemají chloroplastové endoplazmatické retikulum, což je také strukturální rys.
Jak lidé používají červené řasy?
Lidé používají červené řasy jako potravu, protože je známo, že jsou bohaté na jód, vitamíny a bílkoviny. Červené řasy se pěstují za účelem produkce různých prospěšných produktů, které jsou snadno stravitelné a prospěšné lidskému zdraví.
Buňky červených řas obsahují látky, které se používají jako zahušťovadla v cukrářském a potravinářském průmyslu. Lidé se naučili extrahovat tyto látky a používat je k výrobě zmrzliny, marmelády, sýra a omáček. Agaróza a agaropektin se extrahují z buněk řas za vzniku agaru.
Vzhledem k tomu, že některé červené řasy jsou schopny se podílet na tvorbě korálových útesů a zároveň akumulovat látky rozpuštěné ve vodě, lidé je využívají k biologickým a chemickým studiím vody. Červené řasy mohou vykazovat přítomnost chemikálií ve vodě a také záření.
Většina červených řas může být použita jako potrava pro různá zvířata.
















