Pouze dnes: sleva až 20 % jako dárek k vaší první objednávce.
Jakou práci mám napsat?
Struktura mozku kostnatých ryb  Mozek kostnatých ryb se skládá z pěti částí typických pro většinu obratlovců. Diamantový mozek (rhombencephalon) zahrnuje prodlouženou míchu a mozeček. Medulla oblongata (myelencephalon, medulla oblongata) přední část zasahuje pod mozeček a vzadu bez viditelných hranic přechází v míchu. Pro zobrazení přední části prodloužené míchy je nutné otočit tělo mozečku dopředu (u některých ryb je mozeček malý a přední část prodloužené míchy je dobře viditelná). Střechu této části mozku představuje plexus choroideus. Pod ním leží velký kosočtvercová fossa (fossa rhomboidea), rozšířená na předním konci a přecházející vzadu do úzké mediální štěrbiny, je to dutina čtvrtá mozková komora (ventriculus quartus). Medulla oblongata slouží jako počátek většiny mozkových nervů a také jako cesta spojující různá centra předních částí mozku s míchou. Vrstva bílé hmoty pokrývající prodlouženou míchu u ryb je však poměrně tenká, protože tělo a ocas jsou z velké části autonomní – většinu pohybů provádějí reflexně, bez korelace s mozkem. Ve dně medulla oblongata u ryb a ocasatých obojživelníků leží pár obra Mauthnerovy buňky, spojené s akusticko-laterálními centry. Jejich tlusté axony se táhnou podél celé míchy. Pohyb u ryb se provádí hlavně kvůli rytmickému ohýbání těla, které je zjevně řízeno především místními míšními reflexy. Celkovou kontrolu nad těmito pohyby však vykonávají Mauthnerovy buňky. Dýchací centrum leží na dně prodloužené míchy. Při pohledu na mozek zdola můžete rozlišit původ některých nervů. Z boční strany přední části prodloužené míchy vybíhají tři kulaté kořeny. První, ležící nejvíce kraniálně, patří V a VII nervy, střední kořen – pouze VII nerv a konečně třetí kořen, ležící kaudálně, je VIII nerv. Za nimi, také z bočního povrchu prodloužené míchy, se páry IX a X rozkládají společně v několika kořenech. Zbývající nervy jsou tenké a jsou obvykle odříznuty během pitvy. Mozeček Docela dobře vyvinutá, kulatá nebo protáhlá, leží nad přední částí prodloužené míchy přímo za zrakovými laloky. Svým zadním okrajem kryje prodlouženou míchu. Část, která vyčnívá nahoru, je tělo mozečku (corpus cerebelli). Mozeček je centrem pro přesnou regulaci všech motorických inervací spojených s plaváním a uchopováním potravy. Střední mozek (mezencefalon) – část mozkového kmene proniklá mozkovým akvaduktem. Skládá se z velkých, podélně protáhlých optických laloků (jsou viditelné shora). Optické laloky nebo vizuální střecha (lobis opticus s. tectum opticus) – párové útvary oddělené od sebe hlubokou podélnou rýhou. Optické laloky jsou primárními zrakovými centry pro snímání stimulace. V nich končí vlákna zrakového nervu. U ryb má tato část mozku primární význam, je to centrum, které má hlavní vliv na činnost těla. Šedá hmota pokrývající optické laloky má složitou vrstevnatou strukturu, připomínající stavbu mozkové kůry nebo hemisfér.Z ventrálního povrchu optických laloků se rozprostírají silné optické nervy, které procházejí pod povrchem diencefala. Pokud otevřete optické laloky středního mozku, můžete vidět, že v jejich dutině se od mozečku odděluje záhyb, tzv. cerebelární chlopeň (valvule cerebellis). Na obou jeho stranách ve spodní části dutiny středního mozku jsou dvě vyvýšeniny ve tvaru fazole, tzv semilunární tělesa (tori semicircularis) a jsou dalšími centry statoakustického orgánu. přední mozek (prosencephalon) méně vyvinuté než střední, skládá se z telencephalon a diencephalon. Díly diencephalon ležet kolem svislé štěrbiny třetí mozková komora (ventriculus tertius). Boční stěny komory – zrakové hrbolky nebo thalamus (thalamus) u ryb a obojživelníků jsou druhořadé (jako koordinační senzorická a motorická centra). Střecha třetí mozkové komory – epithalamus nebo epithalamus – neobsahuje neurony. Obsahuje přední plexus choroideus (cévní tegmentum třetí komory) a horní dřeňovou žlázu – epifýza (epifíza). Dno třetí mozkové komory – hypotalamu nebo hypotalamu u ryb tvoří párové otoky – dolní laloky (lobus inferior). Před nimi leží spodní dřeňová žláza – hypofýza (hypofýza). U mnoha ryb tato žláza těsně zapadá do zvláštního vybrání na dně lebky a obvykle se během přípravy odlomí; pak jasně viditelné nálevka (infundibulum). Vpředu, na hranici mezi spodní částí terminální a střední části mozku, je optické chiasma (chiasma nervorum opticorum). telencephalon u kostnatých ryb je ve srovnání s jinými částmi mozku velmi malá. Většina ryb (kromě plicník a lalokoploutvých ryb) se vyznačuje evertovanou (převrácenou) strukturou telencephalonových hemisfér. Zdá se, že jsou „otočené“ ventrolaterálně. Střecha předního mozku neobsahuje nervové buňky a skládá se z tenké epiteliální membrány (pallium), který se při pitvě obvykle odstraňuje spolu s membránou mozku. V tomto případě preparát ukazuje dno první komory, rozdělené na dvě hlubokou podélnou rýhou striatum. Striatum (corpora striatum1) sestávají ze dvou částí, které lze vidět při pohledu na mozek ze strany. Ve skutečnosti tyto masivní struktury obsahují striatální a kortikální materiál poměrně složité struktury. Čichové cibule (bulbus olfactorius) přiléhající k přednímu okraji telencefala. Jdou dopředu čichové nervy. U některých ryb (například tresky) jsou čichové bulby umístěny daleko vpředu, v takovém případě se spojují s mozkem čichové dráhy. Kraniální nervy ryb. Z rybího mozku se táhne celkem 10 párů nervů. V podstatě (jak jménem, tak funkcí) odpovídají nervům savců. Struktura žabího mozku Mozku žáby, stejně jako ostatní obojživelníci, se ve srovnání s rybami vyznačují následujícími znaky: a) progresivní vývoj mozku, vyjádřený oddělením párových hemisfér podélnou trhlinou a vývojem šedé hmoty starověké kůry (archipallium) v střecha mozku; b) slabý vývoj mozečku; c) slabé vyjádření ohybů mozku, díky čemuž jsou shora jasně viditelné střední a střední části.
Mozek kostnatých ryb se skládá z pěti částí typických pro většinu obratlovců. Diamantový mozek (rhombencephalon) zahrnuje prodlouženou míchu a mozeček. Medulla oblongata (myelencephalon, medulla oblongata) přední část zasahuje pod mozeček a vzadu bez viditelných hranic přechází v míchu. Pro zobrazení přední části prodloužené míchy je nutné otočit tělo mozečku dopředu (u některých ryb je mozeček malý a přední část prodloužené míchy je dobře viditelná). Střechu této části mozku představuje plexus choroideus. Pod ním leží velký kosočtvercová fossa (fossa rhomboidea), rozšířená na předním konci a přecházející vzadu do úzké mediální štěrbiny, je to dutina čtvrtá mozková komora (ventriculus quartus). Medulla oblongata slouží jako počátek většiny mozkových nervů a také jako cesta spojující různá centra předních částí mozku s míchou. Vrstva bílé hmoty pokrývající prodlouženou míchu u ryb je však poměrně tenká, protože tělo a ocas jsou z velké části autonomní – většinu pohybů provádějí reflexně, bez korelace s mozkem. Ve dně medulla oblongata u ryb a ocasatých obojživelníků leží pár obra Mauthnerovy buňky, spojené s akusticko-laterálními centry. Jejich tlusté axony se táhnou podél celé míchy. Pohyb u ryb se provádí hlavně kvůli rytmickému ohýbání těla, které je zjevně řízeno především místními míšními reflexy. Celkovou kontrolu nad těmito pohyby však vykonávají Mauthnerovy buňky. Dýchací centrum leží na dně prodloužené míchy. Při pohledu na mozek zdola můžete rozlišit původ některých nervů. Z boční strany přední části prodloužené míchy vybíhají tři kulaté kořeny. První, ležící nejvíce kraniálně, patří V a VII nervy, střední kořen – pouze VII nerv a konečně třetí kořen, ležící kaudálně, je VIII nerv. Za nimi, také z bočního povrchu prodloužené míchy, se páry IX a X rozkládají společně v několika kořenech. Zbývající nervy jsou tenké a jsou obvykle odříznuty během pitvy. Mozeček Docela dobře vyvinutá, kulatá nebo protáhlá, leží nad přední částí prodloužené míchy přímo za zrakovými laloky. Svým zadním okrajem kryje prodlouženou míchu. Část, která vyčnívá nahoru, je tělo mozečku (corpus cerebelli). Mozeček je centrem pro přesnou regulaci všech motorických inervací spojených s plaváním a uchopováním potravy. Střední mozek (mezencefalon) – část mozkového kmene proniklá mozkovým akvaduktem. Skládá se z velkých, podélně protáhlých optických laloků (jsou viditelné shora). Optické laloky nebo vizuální střecha (lobis opticus s. tectum opticus) – párové útvary oddělené od sebe hlubokou podélnou rýhou. Optické laloky jsou primárními zrakovými centry pro snímání stimulace. V nich končí vlákna zrakového nervu. U ryb má tato část mozku primární význam, je to centrum, které má hlavní vliv na činnost těla. Šedá hmota pokrývající optické laloky má složitou vrstevnatou strukturu, připomínající stavbu mozkové kůry nebo hemisfér.Z ventrálního povrchu optických laloků se rozprostírají silné optické nervy, které procházejí pod povrchem diencefala. Pokud otevřete optické laloky středního mozku, můžete vidět, že v jejich dutině se od mozečku odděluje záhyb, tzv. cerebelární chlopeň (valvule cerebellis). Na obou jeho stranách ve spodní části dutiny středního mozku jsou dvě vyvýšeniny ve tvaru fazole, tzv semilunární tělesa (tori semicircularis) a jsou dalšími centry statoakustického orgánu. přední mozek (prosencephalon) méně vyvinuté než střední, skládá se z telencephalon a diencephalon. Díly diencephalon ležet kolem svislé štěrbiny třetí mozková komora (ventriculus tertius). Boční stěny komory – zrakové hrbolky nebo thalamus (thalamus) u ryb a obojživelníků jsou druhořadé (jako koordinační senzorická a motorická centra). Střecha třetí mozkové komory – epithalamus nebo epithalamus – neobsahuje neurony. Obsahuje přední plexus choroideus (cévní tegmentum třetí komory) a horní dřeňovou žlázu – epifýza (epifíza). Dno třetí mozkové komory – hypotalamu nebo hypotalamu u ryb tvoří párové otoky – dolní laloky (lobus inferior). Před nimi leží spodní dřeňová žláza – hypofýza (hypofýza). U mnoha ryb tato žláza těsně zapadá do zvláštního vybrání na dně lebky a obvykle se během přípravy odlomí; pak jasně viditelné nálevka (infundibulum). Vpředu, na hranici mezi spodní částí terminální a střední části mozku, je optické chiasma (chiasma nervorum opticorum). telencephalon u kostnatých ryb je ve srovnání s jinými částmi mozku velmi malá. Většina ryb (kromě plicník a lalokoploutvých ryb) se vyznačuje evertovanou (převrácenou) strukturou telencephalonových hemisfér. Zdá se, že jsou „otočené“ ventrolaterálně. Střecha předního mozku neobsahuje nervové buňky a skládá se z tenké epiteliální membrány (pallium), který se při pitvě obvykle odstraňuje spolu s membránou mozku. V tomto případě preparát ukazuje dno první komory, rozdělené na dvě hlubokou podélnou rýhou striatum. Striatum (corpora striatum1) sestávají ze dvou částí, které lze vidět při pohledu na mozek ze strany. Ve skutečnosti tyto masivní struktury obsahují striatální a kortikální materiál poměrně složité struktury. Čichové cibule (bulbus olfactorius) přiléhající k přednímu okraji telencefala. Jdou dopředu čichové nervy. U některých ryb (například tresky) jsou čichové bulby umístěny daleko vpředu, v takovém případě se spojují s mozkem čichové dráhy. Kraniální nervy ryb. Z rybího mozku se táhne celkem 10 párů nervů. V podstatě (jak jménem, tak funkcí) odpovídají nervům savců. Struktura žabího mozku Mozku žáby, stejně jako ostatní obojživelníci, se ve srovnání s rybami vyznačují následujícími znaky: a) progresivní vývoj mozku, vyjádřený oddělením párových hemisfér podélnou trhlinou a vývojem šedé hmoty starověké kůry (archipallium) v střecha mozku; b) slabý vývoj mozečku; c) slabé vyjádření ohybů mozku, díky čemuž jsou shora jasně viditelné střední a střední části. 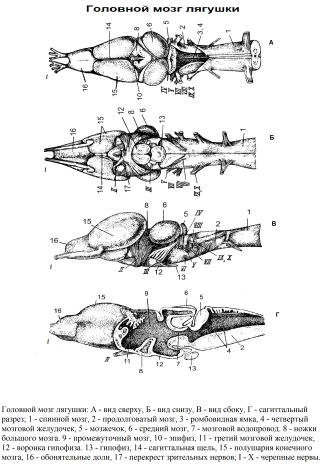 Diamantový mozek (rhombencephalon) Medulla oblongata (myelencephalon, medulla oblongata), do kterého kraniálně přechází mícha, liší se od posledně jmenovaného větší šířkou a odkloněním od svých postranních ploch velkých kořenů zadních hlavových nervů. Na dorzální ploše prodloužené míchy je kosočtvercová fossa (fossa rhomboidea), vstřícný čtvrtá mozková komora (ventriculus quartus). Nahoře je pokryta tenkou cévní uzávěr, který se odstraňuje spolu s mozkovými plenami. Ventrální štěrbina, pokračování ventrální štěrbiny míchy, probíhá podél ventrálního povrchu prodloužené míchy. Medulla oblongata obsahuje dva páry provazců (svazků vláken): spodní pár, oddělený ventrální štěrbinou, je motorický, horní pár je senzorický. Prodloužená dřeň obsahuje centra čelisti a sublingválního aparátu, orgán sluchu a také trávicí a dýchací systém. Mozeček umístěný před kosočtvercovou jamkou v podobě vysokého příčného hřebene jako výrůstek jeho přední stěny. Malá velikost mozečku je dána malou a rovnoměrnou pohyblivostí obojživelníků – ve skutečnosti se skládá ze dvou malých částí, úzce spojených s akustickými centry prodloužené míchy (tyto části jsou u savců zachovány jako fragmenty mozečku (flocculi)). Tělo mozečku, centrum koordinace s ostatními částmi mozku, je velmi špatně vyvinuté. Střední mozek (mezencefalon) při pohledu z dorzální strany je reprezentován dvěma typickými optické laloky (lobus opticus s. tectum opticus), mající vzhled párových vejčitých vyvýšenin tvořících horní a laterální části středního mozku. Střecha optických laloků je tvořena šedou hmotou – několika vrstvami nervových buněk. Tektum u obojživelníků je nejvýznamnější částí mozku. Optické laloky obsahují dutiny, které jsou postranními větvemi cerebrální (Sylvii) akvadukt (aquaeductus cerebri (Sylvii), spojující čtvrtou mozkovou komoru s třetí. Dno středního mozku je tvořeno tlustými svazky nervových vláken – cerebrální stopky (cruri cerebri), spojující přední mozek s prodlouženou míchou a míchou. přední mozek (prosencephalon) sestává z diencephalon a telencephalon, ležící postupně. Diencephalon viditelný shora jako kosočtverec s ostrými úhly směřujícími do stran. Části diencephalonu leží kolem vertikálně umístěné široké trhliny třetí mozková komora (ventriculus tertius). Boční ztluštění stěn komory – zrakové hrbolky nebo thalamus. U ryb a obojživelníků má thalamus druhořadý význam (jako koordinační senzorická a motorická centra). Membranózní střecha třetí mozkové komory – epithalamus nebo epithalamus – neobsahuje neurony. Obsahuje horní medulární žlázu – epifýza (epifíza). U obojživelníků již epifýza slouží jako žláza, ale ještě neztratila rysy parietálního orgánu vidění. Před epifýzou je diencephalon pokryt membranózní stříškou, která se orálně stáčí dovnitř a přechází do předního choroidálního plexu (choroidní tektum třetí komory) a poté do koncové desky diencephala. Dole se komora zužuje, tvoří se nálevka hypofýzy (infundibulum), dolní dřeňová žláza je k ní připojena kaudoventrálně – hypofýza (hypofýza). Vpředu, na hranici mezi spodní částí terminální a střední části mozku, je chiasma nervorum opticorum). U obojživelníků většina vláken zrakových nervů není zadržena v diencefalu, ale jde dále do stropu středního mozku. telencephalon jeho délka je téměř stejná jako délka všech ostatních částí mozku. Skládá se ze dvou částí: čichového mozku a dvou od sebe oddělených hemisfér sagitální (šipovitá) štěrbina (fissura sagittalis). Hemisféry telencephalon (haemispherium cerebri) zaujímají zadní dvě třetiny telencephala a visí přes přední část diencephala, částečně jej zakrývají. Uvnitř hemisfér jsou dutiny – postranní mozkové komory (ventriculi lateralis), kaudálně komunikující s třetí komorou. V šedé hmotě mozkových hemisfér obojživelníků lze rozlišit tři oblasti: dorzomediálně se nachází stará kůra neboli hippocampus (archipallium, s. hippocampus) a laterálně – starověká kůra (paleopallium) a ventrolaterálně – bazální ganglia, odpovídající striata (corpora striata) savců. Striatum a v menší míře hipokampus jsou korelačními centry, posledně jmenovaný je spojen s čichovou funkcí. Starověký kortex je výhradně čichový analyzátor. Na ventrálním povrchu hemisfér jsou patrné rýhy, oddělující striatum od starověkého kortexu. Čichový mozek (rhinencephalon) zaujímá přední část telencephalon a tvoří čichové laloky (cibulky) (lobus olfactorius), pájené uprostřed mezi sebou. Od hemisfér jsou odděleny laterálně marginální jamkou. Čichové laloky vpředu obsahují čichové nervy. 10 párů vyčnívá z mozku žáby lebeční nervy. Jejich tvorba, větvení a zóna inervace se zásadně neliší od těch u savců Ptačí mozek.
Diamantový mozek (rhombencephalon) Medulla oblongata (myelencephalon, medulla oblongata), do kterého kraniálně přechází mícha, liší se od posledně jmenovaného větší šířkou a odkloněním od svých postranních ploch velkých kořenů zadních hlavových nervů. Na dorzální ploše prodloužené míchy je kosočtvercová fossa (fossa rhomboidea), vstřícný čtvrtá mozková komora (ventriculus quartus). Nahoře je pokryta tenkou cévní uzávěr, který se odstraňuje spolu s mozkovými plenami. Ventrální štěrbina, pokračování ventrální štěrbiny míchy, probíhá podél ventrálního povrchu prodloužené míchy. Medulla oblongata obsahuje dva páry provazců (svazků vláken): spodní pár, oddělený ventrální štěrbinou, je motorický, horní pár je senzorický. Prodloužená dřeň obsahuje centra čelisti a sublingválního aparátu, orgán sluchu a také trávicí a dýchací systém. Mozeček umístěný před kosočtvercovou jamkou v podobě vysokého příčného hřebene jako výrůstek jeho přední stěny. Malá velikost mozečku je dána malou a rovnoměrnou pohyblivostí obojživelníků – ve skutečnosti se skládá ze dvou malých částí, úzce spojených s akustickými centry prodloužené míchy (tyto části jsou u savců zachovány jako fragmenty mozečku (flocculi)). Tělo mozečku, centrum koordinace s ostatními částmi mozku, je velmi špatně vyvinuté. Střední mozek (mezencefalon) při pohledu z dorzální strany je reprezentován dvěma typickými optické laloky (lobus opticus s. tectum opticus), mající vzhled párových vejčitých vyvýšenin tvořících horní a laterální části středního mozku. Střecha optických laloků je tvořena šedou hmotou – několika vrstvami nervových buněk. Tektum u obojživelníků je nejvýznamnější částí mozku. Optické laloky obsahují dutiny, které jsou postranními větvemi cerebrální (Sylvii) akvadukt (aquaeductus cerebri (Sylvii), spojující čtvrtou mozkovou komoru s třetí. Dno středního mozku je tvořeno tlustými svazky nervových vláken – cerebrální stopky (cruri cerebri), spojující přední mozek s prodlouženou míchou a míchou. přední mozek (prosencephalon) sestává z diencephalon a telencephalon, ležící postupně. Diencephalon viditelný shora jako kosočtverec s ostrými úhly směřujícími do stran. Části diencephalonu leží kolem vertikálně umístěné široké trhliny třetí mozková komora (ventriculus tertius). Boční ztluštění stěn komory – zrakové hrbolky nebo thalamus. U ryb a obojživelníků má thalamus druhořadý význam (jako koordinační senzorická a motorická centra). Membranózní střecha třetí mozkové komory – epithalamus nebo epithalamus – neobsahuje neurony. Obsahuje horní medulární žlázu – epifýza (epifíza). U obojživelníků již epifýza slouží jako žláza, ale ještě neztratila rysy parietálního orgánu vidění. Před epifýzou je diencephalon pokryt membranózní stříškou, která se orálně stáčí dovnitř a přechází do předního choroidálního plexu (choroidní tektum třetí komory) a poté do koncové desky diencephala. Dole se komora zužuje, tvoří se nálevka hypofýzy (infundibulum), dolní dřeňová žláza je k ní připojena kaudoventrálně – hypofýza (hypofýza). Vpředu, na hranici mezi spodní částí terminální a střední části mozku, je chiasma nervorum opticorum). U obojživelníků většina vláken zrakových nervů není zadržena v diencefalu, ale jde dále do stropu středního mozku. telencephalon jeho délka je téměř stejná jako délka všech ostatních částí mozku. Skládá se ze dvou částí: čichového mozku a dvou od sebe oddělených hemisfér sagitální (šipovitá) štěrbina (fissura sagittalis). Hemisféry telencephalon (haemispherium cerebri) zaujímají zadní dvě třetiny telencephala a visí přes přední část diencephala, částečně jej zakrývají. Uvnitř hemisfér jsou dutiny – postranní mozkové komory (ventriculi lateralis), kaudálně komunikující s třetí komorou. V šedé hmotě mozkových hemisfér obojživelníků lze rozlišit tři oblasti: dorzomediálně se nachází stará kůra neboli hippocampus (archipallium, s. hippocampus) a laterálně – starověká kůra (paleopallium) a ventrolaterálně – bazální ganglia, odpovídající striata (corpora striata) savců. Striatum a v menší míře hipokampus jsou korelačními centry, posledně jmenovaný je spojen s čichovou funkcí. Starověký kortex je výhradně čichový analyzátor. Na ventrálním povrchu hemisfér jsou patrné rýhy, oddělující striatum od starověkého kortexu. Čichový mozek (rhinencephalon) zaujímá přední část telencephalon a tvoří čichové laloky (cibulky) (lobus olfactorius), pájené uprostřed mezi sebou. Od hemisfér jsou odděleny laterálně marginální jamkou. Čichové laloky vpředu obsahují čichové nervy. 10 párů vyčnívá z mozku žáby lebeční nervy. Jejich tvorba, větvení a zóna inervace se zásadně neliší od těch u savců Ptačí mozek.  Diamantový mozek (rhombencephalon) zahrnuje prodlouženou míchu a mozeček. Medulla oblongata (myelencephalon, medulla oblongata) za ním přímo přechází do míchy (medulla spinalis). Vpředu se vklínuje mezi optické laloky středního mozku. Medulla oblongata má tlusté dno, ve kterém leží jádra šedé hmoty – centra mnoha životních funkcí těla (včetně rovnovážně-sluchové, somatické motorické a autonomní). Šedá hmota u ptáků je pokryta silnou vrstvou bílé, tvořenou nervovými vlákny spojujícími mozek s míchou. V dorzální části prodloužené míchy je kosočtvercová fossa (fossa rhomboidea), což je dutina čtvrtá mozková komora (ventriculus quartus). Střechu čtvrté mozkové komory tvoří membranózní cévní tegmentum, u ptáků je zcela pokryto zadní částí mozečku. Mozeček u ptáků je velký a je zastoupen prakticky pouze červ (vermis), nachází se nad prodlouženou míchou. Kůra (šedá hmota umístěná povrchně) má hluboké rýhy, které výrazně zvětšují její plochu. Mozečkové hemisféry jsou špatně vyvinuté. U ptáků jsou části mozečku spojené se svalovým smyslem dobře vyvinuty, zatímco části odpovědné za funkční spojení mozečku s mozkovou kůrou prakticky chybí (vyvíjejí se pouze u savců). Dutina je dobře patrná v podélném řezu cerebelární komora (ventriculus cerebelli), stejně jako střídání bílé a šedé hmoty, tvořící charakteristický vzor strom života (arbor vitae). Střední mozek (mezencefalon) reprezentované dvěma velmi velkými, posunutými do strany zrakové laloky (lobus opticus s. tectum opticus). U všech obratlovců souvisí velikost a vývoj očních laloků s velikostí očí. Jsou dobře viditelné ze strany a z ventrální strany, zatímco z dorzální strany jsou téměř zcela pokryty zadními úseky hemisfér. U ptáků se téměř všechna vlákna očního nervu dostávají do očních laloků a optické laloky zůstávají extrémně důležitými částmi mozku (u ptáků však mozková kůra začíná soutěžit s optickými laloky v důležitosti). Sagitální řez ukazuje, že v dopředném směru dutina čtvrté komory, zužující se, přechází do dutiny středního mozku – mozkový nebo Sylvův akvadukt (aquaeductus cerebri). Orálně akvadukt prochází, expanduje, do dutiny třetí mozkové komory diencephalon. Je vytvořena konvenční přední hranice středního mozku zadní komisura (comissura posterior), jasně viditelné na sagitálním řezu ve formě bílé skvrny. Zahrnuta přední mozek (prosencephalon) existují diencephalon a telencephalon. Diencephalon u ptáků je viditelný zvenčí pouze z ventrální strany. Střední část podélného řezu diencefalem zaujímá úzká vertikální trhlina třetí komora (ventriculus tertius). V horní části komorové dutiny je otvor (párový) vedoucí do dutiny postranní komory – Monroe (interventrikulární) foramen (foramen interventriculare). Boční stěny třetí mozkové komory jsou tvořeny poměrně dobře vyvinutými thalamus, stupeň rozvoje thalamu souvisí se stupněm rozvoje hemisfér. Nemá sice význam vyššího zrakového centra u ptáků, nicméně plní důležité funkce jako motorické korelační centrum. V přední stěně třetí komory leží přední komisura (comissura anterior), skládající se z bílých vláken spojujících obě polokoule.Dno diencefala se nazývá hypotalamus (hypotalamus). Při pohledu zespodu jsou viditelné boční zesílení dna – zrakové dráhy (tractus opticus). Mezi nimi zahrnuje přední konec diencefala zrakové nervy (nervus opticus), formování optické chiasma (chiasma opticum). Zadní dolní roh třetí mozkové komory odpovídá dutině nálevky (infunbulum). Zespodu je nálevka obvykle pokryta podcerebrální žlázou, která je u ptáků dobře vyvinuta – hypofýza (hypofýza). Ze střechy diencephalonu (epitalamus) vybíhající nahoru a mající dutinu stopka epifýzy. Nahoře je on sám šišinkový orgán – epifýza (epifýza), je vidět shora, mezi zadním okrajem mozkových hemisfér a mozečkem. Přední část střechy diencefala tvoří plexus choroideus zasahující do dutiny třetí komory. telencephalon u ptáků se skládá z mozkové hemisféry (hemispherium cerebri), odděleny od sebe hluboko podélná štěrbina (fissura interhemispherica). Hemisféry u ptáků jsou největší útvary mozku, ale jejich struktura se zásadně liší od struktury savců. Na rozdíl od mozku mnoha savců nenesou značně zvětšené hemisféry ptačího mozku rýhy a záhyby, jejich povrch je hladký na ventrální i dorzální straně. Kůra jako celek je špatně vyvinutá, primárně kvůli zmenšení čichového orgánu. Tenkou mediální stěnu hemisféry předního mozku v horní části představuje nervová látka stará kůra (archipallium). Materiál neokortex (špatně vyvinuté) (neopalium) spolu s významnou hmotou striatum (corpus striatum) tvoří silnou boční stěnu hemisféry nebo laterální výrůstek, vyčnívající do dutiny postranní komory. Proto dutina postranní komora (ventriculus lateralis) hemisféra je úzká mezera umístěná dorzomediálně. U ptáků, na rozdíl od savců, se významného vývoje v hemisférách nedosahuje mozková kůra, ale striatum. Bylo odhaleno, že striatum je zodpovědné za vrozené stereotypní behaviorální reakce, zatímco neokortex poskytuje schopnost individuálního učení. Bylo zjištěno, že některé druhy ptáků mají lepší než průměrný vývoj části neokortexu, jako jsou vrány, které jsou známé svými schopnostmi učit se. Cibulky čichové (bulbis olfactorius) nachází se na ventrální straně předního mozku. Jsou malé velikosti a přibližně trojúhelníkového tvaru. Vstupují zepředu čichový nerv.
Diamantový mozek (rhombencephalon) zahrnuje prodlouženou míchu a mozeček. Medulla oblongata (myelencephalon, medulla oblongata) za ním přímo přechází do míchy (medulla spinalis). Vpředu se vklínuje mezi optické laloky středního mozku. Medulla oblongata má tlusté dno, ve kterém leží jádra šedé hmoty – centra mnoha životních funkcí těla (včetně rovnovážně-sluchové, somatické motorické a autonomní). Šedá hmota u ptáků je pokryta silnou vrstvou bílé, tvořenou nervovými vlákny spojujícími mozek s míchou. V dorzální části prodloužené míchy je kosočtvercová fossa (fossa rhomboidea), což je dutina čtvrtá mozková komora (ventriculus quartus). Střechu čtvrté mozkové komory tvoří membranózní cévní tegmentum, u ptáků je zcela pokryto zadní částí mozečku. Mozeček u ptáků je velký a je zastoupen prakticky pouze červ (vermis), nachází se nad prodlouženou míchou. Kůra (šedá hmota umístěná povrchně) má hluboké rýhy, které výrazně zvětšují její plochu. Mozečkové hemisféry jsou špatně vyvinuté. U ptáků jsou části mozečku spojené se svalovým smyslem dobře vyvinuty, zatímco části odpovědné za funkční spojení mozečku s mozkovou kůrou prakticky chybí (vyvíjejí se pouze u savců). Dutina je dobře patrná v podélném řezu cerebelární komora (ventriculus cerebelli), stejně jako střídání bílé a šedé hmoty, tvořící charakteristický vzor strom života (arbor vitae). Střední mozek (mezencefalon) reprezentované dvěma velmi velkými, posunutými do strany zrakové laloky (lobus opticus s. tectum opticus). U všech obratlovců souvisí velikost a vývoj očních laloků s velikostí očí. Jsou dobře viditelné ze strany a z ventrální strany, zatímco z dorzální strany jsou téměř zcela pokryty zadními úseky hemisfér. U ptáků se téměř všechna vlákna očního nervu dostávají do očních laloků a optické laloky zůstávají extrémně důležitými částmi mozku (u ptáků však mozková kůra začíná soutěžit s optickými laloky v důležitosti). Sagitální řez ukazuje, že v dopředném směru dutina čtvrté komory, zužující se, přechází do dutiny středního mozku – mozkový nebo Sylvův akvadukt (aquaeductus cerebri). Orálně akvadukt prochází, expanduje, do dutiny třetí mozkové komory diencephalon. Je vytvořena konvenční přední hranice středního mozku zadní komisura (comissura posterior), jasně viditelné na sagitálním řezu ve formě bílé skvrny. Zahrnuta přední mozek (prosencephalon) existují diencephalon a telencephalon. Diencephalon u ptáků je viditelný zvenčí pouze z ventrální strany. Střední část podélného řezu diencefalem zaujímá úzká vertikální trhlina třetí komora (ventriculus tertius). V horní části komorové dutiny je otvor (párový) vedoucí do dutiny postranní komory – Monroe (interventrikulární) foramen (foramen interventriculare). Boční stěny třetí mozkové komory jsou tvořeny poměrně dobře vyvinutými thalamus, stupeň rozvoje thalamu souvisí se stupněm rozvoje hemisfér. Nemá sice význam vyššího zrakového centra u ptáků, nicméně plní důležité funkce jako motorické korelační centrum. V přední stěně třetí komory leží přední komisura (comissura anterior), skládající se z bílých vláken spojujících obě polokoule.Dno diencefala se nazývá hypotalamus (hypotalamus). Při pohledu zespodu jsou viditelné boční zesílení dna – zrakové dráhy (tractus opticus). Mezi nimi zahrnuje přední konec diencefala zrakové nervy (nervus opticus), formování optické chiasma (chiasma opticum). Zadní dolní roh třetí mozkové komory odpovídá dutině nálevky (infunbulum). Zespodu je nálevka obvykle pokryta podcerebrální žlázou, která je u ptáků dobře vyvinuta – hypofýza (hypofýza). Ze střechy diencephalonu (epitalamus) vybíhající nahoru a mající dutinu stopka epifýzy. Nahoře je on sám šišinkový orgán – epifýza (epifýza), je vidět shora, mezi zadním okrajem mozkových hemisfér a mozečkem. Přední část střechy diencefala tvoří plexus choroideus zasahující do dutiny třetí komory. telencephalon u ptáků se skládá z mozkové hemisféry (hemispherium cerebri), odděleny od sebe hluboko podélná štěrbina (fissura interhemispherica). Hemisféry u ptáků jsou největší útvary mozku, ale jejich struktura se zásadně liší od struktury savců. Na rozdíl od mozku mnoha savců nenesou značně zvětšené hemisféry ptačího mozku rýhy a záhyby, jejich povrch je hladký na ventrální i dorzální straně. Kůra jako celek je špatně vyvinutá, primárně kvůli zmenšení čichového orgánu. Tenkou mediální stěnu hemisféry předního mozku v horní části představuje nervová látka stará kůra (archipallium). Materiál neokortex (špatně vyvinuté) (neopalium) spolu s významnou hmotou striatum (corpus striatum) tvoří silnou boční stěnu hemisféry nebo laterální výrůstek, vyčnívající do dutiny postranní komory. Proto dutina postranní komora (ventriculus lateralis) hemisféra je úzká mezera umístěná dorzomediálně. U ptáků, na rozdíl od savců, se významného vývoje v hemisférách nedosahuje mozková kůra, ale striatum. Bylo odhaleno, že striatum je zodpovědné za vrozené stereotypní behaviorální reakce, zatímco neokortex poskytuje schopnost individuálního učení. Bylo zjištěno, že některé druhy ptáků mají lepší než průměrný vývoj části neokortexu, jako jsou vrány, které jsou známé svými schopnostmi učit se. Cibulky čichové (bulbis olfactorius) nachází se na ventrální straně předního mozku. Jsou malé velikosti a přibližně trojúhelníkového tvaru. Vstupují zepředu čichový nerv.














