Chemické složení červených řas Neorhodomela larix (Otočit) Masuda Peter the Great Bay
Chemické složení červené řasy Neorhodomela larix (Otoč se) Masuda ze zátoky Petra Velikého
Kadnikova I.A., Vishnevskaya T.I., Aminina N.M.
Irina A. Kadniková, Tatiana I. Višněvskaja, Natalia M. Aminina
Pacifické výzkumné středisko pro rybolov –
TINRO-Center (Vladivostok, Rusko)
červené řasy Neorhodomela larix Z hlediska chemického složení je řád Ceramiales typickým zástupcem oddělení Rhodophyta. Bylo stanoveno jeho makro- a mikroprvkové složení, včetně toxických prvků. Byly provedeny studie složení aminokyselin proteinu a složení mastných kyselin lipidů v modřínu Neorhodomela.
Klíčová slova: červené řasy; neorodomela; chemické složení; kompozice aminokyselin, mastných kyselin a minerálních látek; toxické prvky.
Zátoka Petra Velikého se vyznačuje významnou druhovou diverzitou červených řas (Perstenko, 1994). Mezi potenciálně komerční druhy patří zástupci řádu Ceramiales, rodiny Rhodomelaceae, jehož typový rod je Neorhodomela (Neorhodomela Masuda). Rod Neorhodomela je v mořích Dálného východu zastoupena sedmi druhy, ale rozšířeným druhem je Neorhodomela Larix.
Proto byl jako objekt této studie zvolen nejhojnější zástupce rodu., žijící u pobřeží Přímořského kraje – Neorhodomela Larix (Otočte) Masuda (modřín neohodomelský).
Tato řasa roste v litorálních a sublitorálních zónách do hloubky 3–5 m, na kamenitých a kamenitých půdách, vytváří malé shluky nebo doprovází houštiny řas chaluh. Thallus Neorhodomela je huňatý, rozvětvený, pružný, chrupavčitý, 5–15, někdy až 50 cm vysoký, tmavě hnědé barvy. Větvení stélky je bohaté, v hlavních větvích nepravidelně střídavé, místy fascikulované. Axiální výhon a boční větve jsou vejčité, 1,0–1,5 mm silné. Větve 4.–5. řádu jsou pokryty jednoduchými nebo složitými ostnatými větvemi a větvemi omezeného růstu, uspořádanými rovnoměrně spirálovitě. Řasa je připevněna malou podrážkou, ze které se vyvíjí několik výhonků. Maximální biomasa Neorhodomela může dosáhnout 7,5 kg/m2. U pobřeží Primorye roste po celý rok (Blinova, 1971; Klochkova, 1996; Atlas hromadných druhů řas. 2008).
Modřín neorhodomelský je na ruském Dálném východě rozšířen ve všech pobřežních oblastech a lze jej použít jako surovinu pro výrobu gelotvorných látek. Zásoby modřínu neohodomelského v celém rozsahu dosahují podle odborných odhadů 100–150 tisíc tun (Sukhoveeva, Podkorytova, 2006).
V souvislosti s masivním rozvojem tohoto málo probádaného druhu řasy se jeví jako relevantní studovat její chemické složení a nutriční hodnotu.
Účelem této práce je studovat chemické složení druhu řas, které jsou perspektivní pro rybolov a rostou v pobřežních oblastech Přímořského kraje.
Materiály a metody výzkumu
Jako předmět studia byly použity červené řasy Neorhodomela Larix od rodiny Rhodomelaceae, těžené v zátoce Petra Velkého v červnu 2016.
Celkový obsah minerálních látek v řasách byl stanoven podle GOST 26185-84.
Příprava vzorku pro stanovení atomové absorpce toxických kovů byla provedena podle GOST 26929-94. Obsah makro- a mikroprvků (včetně toxických) v řasách byl stanoven pomocí plamenového emisního spektrofotometru Nippon Jarrell Ash, model AA-855; As a Cd byly stanoveny v grafitové cele na atomovém absorpčním spektrofotometru Shimadzu AA-6800.
Celkový obsah dusíkatých látek byl stanoven Kjeldahlovou mikrometodou pomocí Kjeltec auto 10 SO Analyzer (Tecator, Japonsko). Příprava vzorku pro analýzu aminokyselinového složení proteinu byla provedena metodou kyselé hydrolýzy řasy 6N HCI (Baratova, Belyakova, 1974; Osterman, 1985). Aminokyselinové složení proteinu bylo stanoveno pomocí automatického analyzátoru aminokyselin L-8800 (Hitachi, Japonsko).
Celkový obsah lipidů byl stanoven metodou Blight–Dyer (Blight and Dayer, 1959). Složení mastných kyselin v lipidech bylo stanoveno ve formě methylesterů mastných kyselin (Carreau, Dubacq, 1978) na plynokapalinovém chromatografu GC-16 A (Shimadzu, Japonsko), s plamenově ionizačním detektorem, kapilární kolonou o střední polarita (Carbowax-20) v proudu helia 28 cm/s, teplota injektoru a detektoru 240°С, termostat – 190°С, doba analýzy – 60 min. Mastné kyseliny byly identifikovány pomocí retenčních indexů mastných kyselin, které byly stanoveny individuálně pro každou mastnou kyselinu.
Obsah snadno hydrolyzovatelných polysacharidů byl stanoven kolorimetrickou metodou při vlnové délce 620 nm (Krylova a Lyaskovskaya, 1965).
Výsledky a jejich diskuse
Výsledky chemické analýzy Neorhodomela jsou uvedeny na Obr. 1.

Poznámka: *LGP – snadno hydrolyzovatelné polysacharidy
Rýže. 1. Chemické složení neorhodomely (v % sušiny)
Obr. 1. Chemické složení Neorhodomela larix (% na sušinu)
U modřínu Neorhodomela byly minerální látky stanoveny v množství 30,3 %, sacharidy – 26 %, včetně snadno hydrolyzovatelných polysacharidů a vlákniny, bílkoviny – 16,7 %. Obsah lipidů je na úrovni 1,0 %, což je srovnatelné s údaji pro tento ukazatel pro řasy čeledi Rhodomelaceae, Zátoka Petra Velikého (Khotimchenko, Gusarova, 2004). Tato řasa se vyznačuje velmi nízkým obsahem jódu (0,005 %). Analýza chemického složení této řasy ukázala její podobnost s chemickým složením СHondrus Crispus и Meristotheca papilosa, které jsou zdroji karagenanu (Podkorytova et al., 1994; Chapman, Chapman, 1980).
Mezi biogenními prvky byly identifikovány čtyři makroprvky (K, Na, Ca a Mg) a pět mikroprvků (Mn, Fe, Zn, Cu, Ni). Srovnávací analýza makro- a mikroprvků modřínu Neorhodomela z pobřežních vod obce. Hassan zjistil maximální koncentrace K (54011,3 mg/kg), Na (35369,5 mg/kg), Ca (8530,3 mg/kg), Mg (4993,3 mg/kg) a Zn (58,3 mg/kg) a minimální – Mn (29,9 mg/kg) a Cu (5,0 mg/kg), ve srovnání s komerčními řasami Ahnfeltia tobuchiensis (Kadníková, 2011). Převládající makroživinou v Neorhodomele je K, stejně jako u hnědých řas saccharina japonská (Aminina a kol., 2007). Mezi mikroelementy v tkáních neorhodomel převažuje Fe a Zn (tab. 1).
Tabulka 1. Elementární složení červených řas Neorhodomela Larix (mg/kg suchých řas)
Tabulka 1. Elementární složení červené řasy Neorhodomela larix (mg/kg suchých řas)

Analýza ukázala, že studované druhy řas akumulují více monovalentních kationtů (draslíku и sodík) než dvojmocný (vápníku и hořčíku) a podobá se například dalším zástupcům červených řas Ralmaria palmata (Linnaeus) Weber et Mohr (Rødde a kol., 2004) a СHondrus armatus (Harvey) Okamura (Podkorytova et al., 1994).
Posouzení obsahu toxických prvků modřínu neorhodomelského z vodní plochy obce. Khasan prokázal nesoulad s požadavky TR CU 021/2011 (Technické předpisy. 2011) z hlediska množství As, jehož koncentrace v řasách překračuje maximální přípustnou úroveň (MAL) 6,7krát ( 5,0 mg/kg). Obsah Pb se blíží maximálnímu přípustnému limitu (0,5 mg/kg) a Cd a Hg jsou výrazně nižší než normované hodnoty (

Rýže. 2. Obsah toxických prvků v neorodomelu (mg/kg syrové řasy)
Obr. 2. Obsah toxických prvků v Neorhodomela larix (mg/kg syrových řas)
Tabulka 2 ukazuje obecné složení aminokyselin proteinu neorodomela. Bylo zjištěno, že tato řasa obsahuje 17 aminokyselin typických pro červené řasy, včetně 7 esenciálních (tryptofan nebyl stanoven).
Tabulka 2. Obsah aminokyselin v proteinu Neorhodomela (g/100 g proteinu)
Tabulka 2. Obsah aminokyselin v Neorhodomela larix bílkoviny (g/100 g bílkovin)

Biologická hodnota proteinu Neorhodomela byla hodnocena podle obsahu esenciálních aminokyselin (EAA). Obsah NAC je na úrovni 31,9 % z celkového množství aminokyselin. Množství lysinu a threoninu je vyšší než standardy doporučené FAO/WHO o 24,7 %, resp. 27,5 % (FAO/WHO, 1991) a obsah valinu, leucinu a isoleucinu v proteinech je o něco nižší (tabulka 2). . Neorhodomela se od komerční červené řasy Ahnfeltia tobuchinskaya liší vyšším obsahem esenciálních aminokyselin (EAA), jako je lysin, threonin, isoleucin, leucin, fenylalanin, valin a methionin (Kadniková et al., 2015).
Z neesenciálních aminokyselin (EAA) převládají monoaminodikarboxylové kyseliny.: kyseliny glutamové a asparagové. Obsah prolinu a histidinu v proteinu Neorhodomela je vyšší než u Ahnfeltia a množství ZAC je nižší (Kadniková et al., 2015).
Jednou z hlavních identifikačních charakteristik lipidů je jejich složení mastných kyselin (MK), které je uvedeno v tabulce 3.
Tabulka 3. Složení mastných kyselin z celkových lipidů Neorhodomela (% celkových mastných kyselin)
Tabulka 3. Složení mastných kyselin z celkových lipidů v Neorhodomela larix (% součtu mastných kyselin)

Nasycené mastné kyseliny (SFA) v neorodomelu tvoří 77,0 %, hlavní část tvoří kyselina myristová (14:0) a palmitové (16:0) (10,1 a 59,1 %). Podíl mononenasycených mastných kyselin (MUFA) v neorodomelu je na úrovni 19,1 %, z toho množství kyseliny palmitolejové (16:1 n-7) je téměř 3krát vyšší než u Ahnfeltia Tobuchinskaya (Khotimchenko, 2003). Množství polynenasycených mastných kyselin (PUFA) v neorodomelu je 3,6 %, což je v průměru 2,0krát méně než u ahnfeltia. Poměr n-6 ku n-3 PUFA v neorhodomelštině ze studované oblasti je 3,2:1.
Získané výsledky o kvalitě lipidů Neorhodomela Larix jsou v souladu s literárními údaji o závislosti složení mastných kyselin na druzích makrofyt (Khotimchenko, 2003).
Závěry
Byl proveden výzkum chemického složení červených řas řádu Ceramiales, rostoucí v pobřežní zóně Primorye. Protein v neorodomelu obsahuje 16,7 % sušiny, snadno hydrolyzovatelné sacharidy tvoří 17,2 %. Studované řasy jsou bohaté na makroprvky, jako je K a Na. Mezi mikroprvky převažuje Fe a Zn. Koncentrace toxických prvků v Neorhodomela se liší: hladina Cd a Hg je výrazně nižší než normované hodnoty, hodnota As je vyšší a Pb je na úrovni normy stanovené pro řasy platnými technickými předpisy. Množství NAC v neorodomelu překračuje adekvátní úroveň denní spotřeby rostlinných bílkovin, regulovanou MP 2.3.1. 1915-04 (Doporučené úrovně příjmu. 2004). Obsah SFA v neorodomelu je 77,0 %, MUFA – 19,1 %, PUFA – 3,6 % z celkových MK. S přihlédnutím k chemickému složení lze studovaný typ řasy doporučit pro výrobu polysacharidů, ale i technických produktů (krmiva, přísady do krmiv, hnojiva).
1. Aminina N.M., Vishnevskaya T.I., Guruleva O.N., Kovekovdova L.T. Složení a možnosti využití hnědých řas moří Dálného východu // Bulletin pobočky Dálného východu Ruské akademie věd. 2007. č. 6. s. 123–130.
2. Atlas hromadných druhů řas a mořských trav ruského Dálného východu / Dzizyurov V.D., Kulepanov V.N., Shaposhnikova T.V., Sukhoveeva M.V., Gusarova I.S., Ivanova N.V.; TINRO-Center – Vladivostok: TINRO-Center, 2008. – 327 s.
3. Baratova L.A., Belyakova L.P. Stanovení aminokyselinového složení proteinů // Metody biochemického experimentu. – M., 1974. – S. 1–36.
4. Blínová E.I. Hlavní řasy severovýchodní části Okhotského moře (distribuce, ekologie, fytomasa) // Rostlinné zdroje. 1971. T.7. s. 252–259.
5. GOST 26185-84 Mořské řasy, mořské trávy a produkty jejich zpracování. Metody analýzy. – M.: Standard, 1984. – 53 s.
6. GOST 26929-94 Suroviny a potravinářské výrobky. příprava vzorků Mineralizace pro stanovení toxických prvků. – M., 1994. – 123 s.
7. Kadniková I.A. Technochemické vlastnosti mořských rostlinných surovin pro výrobu gelotvorných polysacharidů // Skladování a zpracování zemědělských surovin. 2011. č. 9. s. 26–29.
8. Kadniková I.A., Aminina N.M., Mokretsova N.D., Rogov A.M. Použití různých druhů řas ve složení krmiva pro mladé mořské okurky // Bulletin ASTU. 2015. č. 4. s. 62–68.
9. Klochková N.G. Flóra řas-makrofytů Tatarského průlivu (Japonské moře) a rysy jeho formování. – Vladivostok: Dalnauka, 1996. – 290 s.
10. Krylova N.N., Lyaskovskaya Yu.N. Fyzikálně-chemické metody studia produktů živočišného původu – M.: Food Industry, 1965. – S. 34–38.
11. MR 2.3.1.1915-04. Doporučené úrovně spotřeby potravin a biologicky aktivních látek – M., 2004. – 41 s.
12. Osterman L.A. Chromatografie proteinů a nukleových kyselin. – M., 1985. – 536 s.
13. Perestenko L.P. Červené řasy Dálného východu moří Ruska. – Petrohrad, 1994. – 331 s.
14. Podkorytová A.V., Kadníková I.A., Úsov A.I. červené řasy Chondrus armatus (Harv.) Dobře. (Gigartinaceae), jeho chemické složení, obsah karagenanu // Rostlinné zdroje. 1994. Číslo 1–2. s. 79–85.
15. Suchoveeva M.V., Podkorytova A.V. Komerční řasy a byliny moří Dálného východu: biologie, distribuce, zásoby, technologie zpracování. – Vladivostok: TINRO-Center, 2006. – 243 s.
16. Technické předpisy Celní unie „O bezpečnosti potravin. Technické předpisy celní unie“, TR CU 021/2011 Schváleno Rozhodnutím Komise celní unie ze dne 09. prosince 2011 č. 880. – Petrohrad: GIORD, 2015. – 242 s.
17. Chotimčenko S.V. Lipidy mořských řas, makrofyt a bylin: Struktura, distribuce, analýza. – Vladivostok: Dalnauka, 2003. – 234 s.
18. Khotimchenko S.V., Gusarova I.S. Červené řasy zálivu Petra Velikého jako zdroj kyseliny arachidonové a eikosapentaenové // Biologie moře. 2004. T.30. č. 3. s. 215–218.
19. Blight E.G., Dayer W.J. Rychlá metoda celkové extrakce lipidů // Kanada. J. Biochem. Physiol. 1959. č. 37. R. 911–917.
20. Carreau J.P., Dubacq J.P. Přizpůsobení metody v makroměřítku na mikroměřítko pro transesterifikaci metylesteru mastných kyselin biologických lipidových extraktů // J. Chromatogr. 1978. V.151. č. 3. S. 384–390.
21. Chapman V.J., Chapman D.J. Mořské řasy a jejich využití. – 3. Londýn: New York, 1980. – 334 s.
22. FAO/WHO, Hodnocení kvality bílkovin (Zpráva o společném vývozním závěru FAO/WHO). – FAO Řím, 1991.
23. Rødde R.S.H., Vårum K.M., Larsen B.A., Myklestad S.M. Sezónní a geografické rozdíly v chemickém složení červených řas Palmaria palmata (L.) Kuntze. // Botanica Marina. 2004. V.47. S. 125–133.
Článek obdržela redakce 1.12.2017. května XNUMX
Chemické složení červené řasy Neorhodomela larix (Otočit) Masuda
ze zátoky Petra Velikého
Irina A. Kadniková, Tatiana I. Višněvskaja, Natalia M. Aminina
Pacific Scientific Research Fisheries Center – TINRO-Сentre (Vladivostok, Rusko)
Chemickým složením červená řasa Neorhodomela larix řád Ceramiales je typickým zástupcem divize Rhodophyta. Bylo odhaleno jeho makro- a mikroprvkové složení včetně toxických prvků. Výzkum proteinových aminokyselin a složení lipidových mastných kyselin Neorhodomela byly provedeny.
Klíčová slova: červené řasy; Neorhodomela; chemické složení; aminokyselina; kompozice mastných kyselin a minerálních látek; toxické prvky.
O autorech
Kadniková Irina Arnoldovna
Doktor technických věd
Vedoucí výzkumný pracovník, Laboratoř problémů racionálního využívání řas, Federal State Unitary Enterprise TINRO-Center (FSUI „Pacific Scientific Research Fisheries Center“ – „TINRO-centrum“)
Višněvská Taťána Ivanovna
kandidát technických věd
Vedoucí výzkumný pracovník, Laboratoř problémů racionálního využívání řas, Federal State Unitary Enterprise TINRO-Center (FSUI „Pacific Scientific Research Fisheries Center“ – „TINRO-centrum“)
Aminina Natalia Michajlovna
kandidát biologických věd
Vedoucí laboratoře problémů racionálního využívání řas ve federálním státním jednotném podniku TINRO-Center (FSUI „Pacific Scientific Research Fisheries Center“ – „TINRO-centrum“)
Korespondentská adresa: Rusko, 690091, Vladivostok, pruh. Shevchenko, 4, FSUE TINRO-centrum; telefon (423) 240-13-60
ODKAZ NA ČLÁNEK:
Kadnikova I.A., Vishnevskaya T.I., Aminina N.M. Chemické složení červených řas Neorhodomela larix (Turn) Masuda ze zátoky Petra Velikého // Problematika moderní algologie. 2018. č. 1 (16). URL: http://algology.ru/1249
Drazí kolegové! Pokud byste chtěli získat PDF verzi článku, napište nám do redakce a my vám ji rádi zdarma zašleme.
Adresa — info@algology.ru
Při opětovném tisku je vyžadován odkaz na web
Už jste někdy viděli kvetoucí jezírko? Ten samý svůdný rybník v horkém červencovém dni, ve kterém se tak chcete koupat. Když se k ní ale přiblížíte, touha často zmizí – voda je zelená, kalná, navíc páchne bažinou. Kdo může za takové zklamání? Odpověď je jednoduchá: řasy! Budeme o nich mluvit v tomto článku.
Obecná charakteristika řas
řasy – skupina nižších rostlin včetně mnoha divizí. Ve školních osnovách se podrobně studují:
Rhizoidy – jednobuněčné orgány přichycení k substrátu. Jsou to zvláštní analogy kořenů, které jsou vlastní organismům, které je nemají.
Proč jsou rhizoidy horší než kořeny?
Hlavní nevýhodou rhizoidů je, že jsou jednobuněčné. Když se pozorně podíváme na strukturu kořene, uvidíme tam mnoho zón, tkání a dalších struktur. Kořen se tedy podobá silnému vodnímu potrubí, kterým do rostliny vstupují živiny. Rhizoidy jsou zastoupeny pouze jednou buňkou, jsou velmi malé a slabé, takže nemohou plnit funkci pohlcování látek. Jsou, stejně jako stopy, potřebné pouze k tomu, aby zabránily tomu, aby byla rostlina unesena proudem. S čím se úspěšně vyrovnávají.
Tělo jakékoli řasy představuje stélka, popř stélka. Může být jednobuněčný nebo mnohobuněčný, různých tvarů.
Thallus – jediná buňka nebo jejich nediferencovaný shluk, který tvoří tělo organismu.
“Nediferencované“ znamená „identické“, to znamená, že všechny buňky v talu mají podobnou strukturu a vykonávají stejné funkce. Podobně jako tužky: liší se pouze barvou a všechny jsou potřebné ke kreslení.
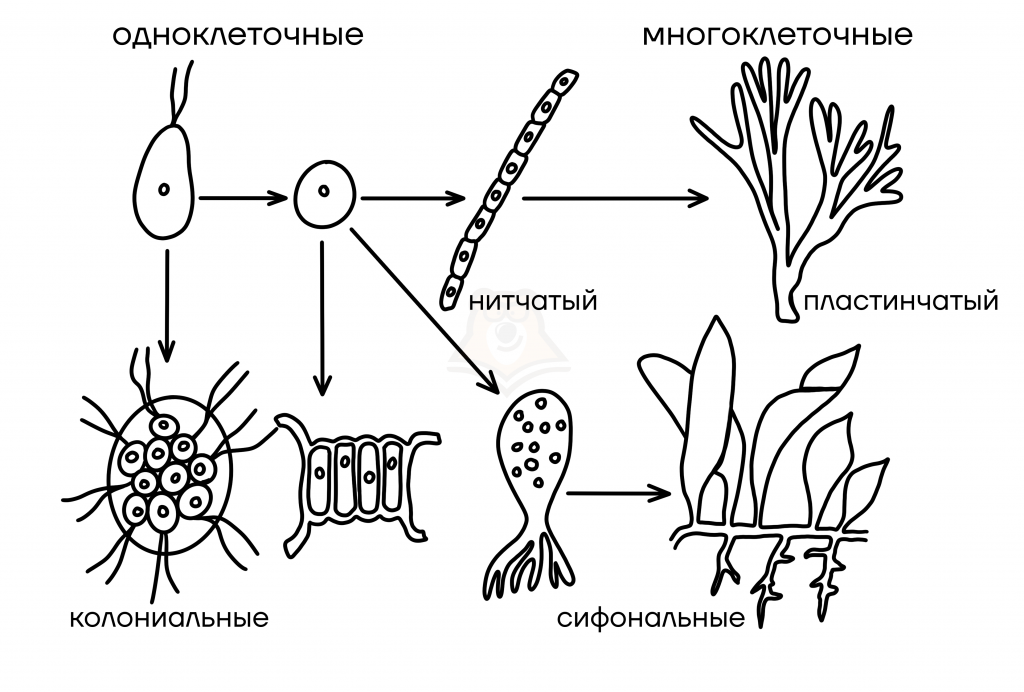
Typy organizace thalli
Typy organizace jednobuněčných stélků
- Monadický – bičíkový. Pro zapamatování termínu si připomeneme zástupce tohoto typu – Chlamydomonas. Písmeno „X“ ve svém názvu připomíná dva bičíky.
- Coccoid – kulový. S kořenovým „kokem“ se setkáme i při studiu mikrobiologie: koky jsou bakterie, které připomínají kuličky, například streptokok. Z tohoto důvodu si pamatujme, že písmeno „o“ ve slově „coccus“ je kulaté jako míč.
- Améboid – nemají stálý tvar. Slovo pochází z názvu nejjednoduššího zvířete – améby, která vypadá jako skvrna. Zhruba takto vypadá stélka améboidních řas.
Typy organizace mnohobuněčných stélků
- vláknitý – reprezentované několika buňkami spojenými v dlouhém řetězci. Navenek připomíná nit, proto dostal své jméno.
- Lamelární – skládající se z několika nití propojených do „desky“. Takové řasy jsou nám nejznámější: vzhledem připomínají vyšší rostlinu s listy.
- Sifon Typ organizace talu je reprezentován mnoha fúzovanými buňkami. Uvnitř takové řasy nezůstaly žádné přepážky, je to jakýsi „rosol“ z cytoplazmy všech buněk.
- sifonokladský stélka se také skládá ze srostlých buněk, ale stále obsahuje nějaké přepážky. „Poklad“ zní, jako by bylo něco někde umístěno. Připomeňme si toto: v sifonokladickém talu leží buněčné struktury uspořádaněji, jako by byly rozloženy na policích.
Ekologie a význam
Řasy žijí nejen ve sladkých a slaných vodách – lze je najít i ve vzorku vody z louže v parku nebo v půdě. Některé druhy se nacházejí v ledu za polárním kruhem. Když se nad tím zamyslíte: řasy jsou nezničitelné jako starý telefon Nokia. I když se nám zdá, že řasy někde nežijí, s největší pravděpodobností jsme na omylu.
V ekosystémech hrají roli řasy výrobci a tvoří potravu pro vodní živočichy.
Řasy mohou být volně žijící nebo součástí lišejníků. Lišejníky – symbiotické organismy, jejichž těla jsou složena z jednotlivých buněk řas umístěných mezi houbovými hyfami. O tom, co to je hyfy Více se dočtete v článku „Houby. Lišejníky.”
Využití řas lidmi
Někteří zástupci (například chaluha – chaluha, ulva – mořský salát) lidé odedávna využívali jako potravu. Asijské kuchyně jsou proslulé především pokrmy z mořských řas. Například Japonci svou láskou k salátu chuka nakazili celý svět: dokonce se dá sehnat i v některých ruských supermarketech.
Červené řasy používá se v průmyslu k výrobě želatinových látek, například agar-agaru.
Agar-agar – směs polysacharidů používaná k přípravě různých kulinářských pokrmů nebo substrát, na kterém v laboratoři dobře rostou kolonie různých organismů.
Nyní přejděme k oddělením řas.
Zelené řasy
Oddělení zelených řas je tak pojmenováno kvůli charakteristickému zbarvení svých zástupců, způsobeném převahou chlorofylu a a b nad ostatními pigmenty.
zástupci: chlamydomonas, chlorella, chlorococcus, ulotrix, acetabularia, volvox, ulva.
Jinakost
Chlorofyl je rostlinný pigment nezbytný pro fotosyntézu. Různé skupiny řas mají různé složení pigmentu. Pigmenty jsou barviva, která určují barvu rostliny.
Škrob je uložen v zelených řasách v pyrenoidu, zvláštním místě v buňce, které obsahuje mnoho enzymů. Tyto enzymy k sobě doslova přitahují molekuly glukózy a spojují je do jednoho velkého škrobového zrna.
- Převládající generace je haploidní.
To znamená, že existuje jedna sada chromozomů. Dospělou rostlinu jakékoli zelené řasy představuje haploidní generace – gametofyt.
| Připomeňme si pravidlo tří “G” – bude se nám nejednou hodit při studiu botaniky. Gametofyt – gamety – haploid. Všechna tato slova začínají písmenem „G“, což znamená, že jsou propojena: gametofyt a gamety mají haploidní sadu chromozomů, gamety se tvoří z gametofytu. |
Nejvýznamnější představitelé odd
Chlamydomonas – jednobuněčná řasa s monadickým stélkem.

V přední části buňky jsou 2 bičíky. Fotosyntetický aparát představuje velký pohárkovitý chromatofor, vyplňující téměř celou cytoplazmu buňky. Škrobová zrna se hromadí v centrální oblasti chromatoforu – pyrenoidu. Existuje fotosenzitivní organela – stigma.
Řasy se rozmnožují nepohlavně i pohlavně.
Během nepohlavní reprodukce se cytoplazma uvnitř buňky mnohokrát dělí mitózou, čímž dochází k porušení celulózové membrány. Ukazuje se, že dospělá řasa nikdy nepřežije vzhled „dětí“.

Zoospora – výtrus, který má jeden nebo více bičíků. Vzpomeňme, kde jinde jsme se setkali s kořenem „zoo“: například ve slovech „zoologie“ (nauka o zvířatech), „zoo“. Zoospora vypadá jako malé zvíře: aktivně se pohybuje a má jakýsi „ocásek“.
Chlorella, pleurokok – jednobuněčná řasa s kokoidním stélkem.
Tyto řasy se rozmnožují pouze nepohlavně s tvorbou bičíkatých autospor.
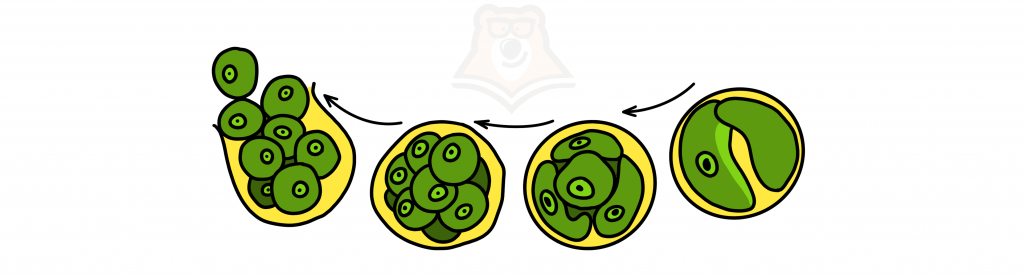
Autospora – nepohyblivá bičíková výtrus.
Pojem si můžeme zapamatovat takto: autosport je AUTONOMNÍ a nezávislý, tento svět je mu již naprosto jasný. Proto tiše leží a nehýbe se, ale přemítá o tom, co se kolem ní děje.

Autospor poznal smysl existence
Pleurococcus často pokrývá kmeny stromů, jeho akumulace vypadají jako zelený povlak na kůře.
Acetabularia (“sklo mořské panny”) je velká mnohobuněčná řasa se sifonovým stélkem.
Zajímavé je, že v celém těle acetabularia je pouze jedno jádro. Právě to řídí životně důležité procesy probíhající v tak velkém organismu.
Volvox – kulovitý koloniální organismus skládající se z mnoha bičíkatých buněk.

Struktura kolonie Volvox
Kolonie – specifická forma soužití jednobuněčných organismů. Buňky v kolonii jsou na sobě nezávislé a mohou existovat samostatně.
Volvox se rozmnožuje nepohlavně (dělením kolonie) nebo pohlavně (s tvorbou bičíkových gamet).
Červené řasy – fialové řasy
Červené řasy – oddělení, jehož představitelé se vyznačují absencí bičíkového stadia v životním cyklu.
zástupci: phyllophora, rhodémie, anfelcia, delesseria.
Jinakost
- Speciální pigmenty: fykoerythrin a fykocyanin.
- Rezervní látkou je purpurový škrob.
- Všechny fáze životního cyklu jsou nehybné.

Červené řasy žijí hlavně v mořských vodách, ve velkých hloubkách ve srovnání s jinými řasami. Fialové rostliny jsou mnohem více přizpůsobeny životu v hloubce než jiné nižší rostliny. Jde o to, že mají červené a modré pigmenty (fykoerythrin a fykocyanin), což jim umožňuje využívat energii slunečních paprsků v modré části spektra pro fotosyntézu. A jen takové paprsky pronikají do velkých hloubek. Takhle šarlatové dívky přelstily systém.

Fialové rostliny obešly fotosyntézu
Hnědé řasy
zástupci: řasa, sargassum, fucus, cystoseira.
Jinakost
- Speciálním pigmentem je fukoxanthin.
- Rezervní látkou je laminarin.
- Většinu svého životního cyklu prožívají v diploidní fázi.
Thalli z hnědých řas mají nejvíce složitá struktura mezi všemi zástupci řas.
Tvoří se tkáňové struktury například sítové buňky. Stále je však nelze nazývat skutečnými tkáněmi, protože všechny buňky mají stejný původ. Podle některých vědců to byly hnědé řasy, které daly vzniknout moderním vyšším rostlinám.

Chaluha – velká mnohobuněčná řasa s lamelárním stélkem. Používané lidmi k jídlu.
Laminaria je známá svým vysokým obsahem jod, což je zvláště užitečné pro lidi s hypotyreózou. Rozmnožuje se pohlavně i nepohlavně, gamety a spory mají bičíky.
Kontrola faktů
- řasy – skupina nižších rostlin včetně mnoha divizí.
- Představené tělo stélkak přichycení k substrátu dochází v důsledku rhizoidů.
- Oddělení zelené řasy tak pojmenován kvůli charakteristickému zbarvení zástupců, způsobeném převahou chlorofylů a a b nad ostatními pigmenty. Hromadí škrob jako rezervní látku a většinu svého životního cyklu žijí v haploidní fázi.
- Červené řasy – oddělení, jehož představitelé se vyznačují absencí bičíkového stadia v životním cyklu.
- Hnědé řasy Vyznačují se ukládáním polysacharidu laminarinu v buňkách a přítomností speciálního pigmentu fukoxanthinu.
zkontroluj se
1 úloha.
Proč jsou řasy považovány za nižší rostliny?
- kvůli malému počtu
- kvůli nedostatku různých forem
- protože nemají skutečné tkáně a orgány
- protože mají příliš malou velikost těla
2 úloha.
Jakou rezervní látku mají hnědé řasy?
3 úloha.
Proč šarlatové ryby žijí ve velkých hloubkách?
- mají červené a modré pigmenty a mohou zachytit paprsky modré části spektra
- připevněte pouze na speciální skalnatý substrát
- žijí v symbióze s hlubokomořskými živočichy
- díky velké tělesné hmotě nevyplavou na hladinu
4 úloha.
Laminaria je užitečná pro lidi s…
- сахарным диабетом
- diabetes insipidus
- hypofunkce štítné žlázy
- hyperfunkce štítné žlázy
5 úloha.
- fotosyntetické zařízení
- hromadění enzymů
- chromatofor
- fotosenzitivní organoid
Odpovědi: 1 – 3; 2 – 4; 3 – 1; 4 – 3; 5 – 2.
















